
Developmental Biology
Introduction
Research has focused on the importance and function of cell-specific transcription factors in the differentiation of cells and in embryogenesis. Of particular interest here are transcription factors whose malfunction leads to diseases such as cancer, diabetes and kidney failure in humans. Cell cultures were used as experimental systems to study cell differentiation and embryos of the Xenopus clawed frog were used as a model for development.
Transgenic Xenopus strains
DNA CONSTRUCTS
The most important DNA constructs produced in the laboratory are available from addgene: https://www.addgene.org/Gerhart_Ryffel/
CELL LINES
INS-1 and HEK293 cell lines for conditional expression of transgenes as published (Thomas et al. 2004, Lucas et al. 2005, Senkel et al. 2005, Erdmann et al. 2007, Senkel et al. 2009, Waldner et al. 2011, submitted).
TRANSGENIC STRAINS OF XENOPUS LAEVIS
Deposited at the European Xenopus Resource Centre of the University of Portsmouth http://www.port.ac.uk/research/exrc and at the Marine Biological Laboratory (MBL), Woods Hole http:/www.mbl.edu/xenopus/laevis-stocks
(a) REPORTER STRAINS
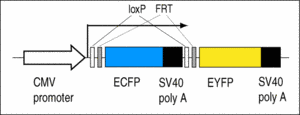
C5: Contains the Cre/FLP reporter LCMV:ECFP(loxP)(FRT)EYFP.
The larvae show blue fluorescence, which changes to yellow fluorescence when exposed to Cre or FLP recombinase (Ryffel et al., 2003; Waldner et al., 2006).
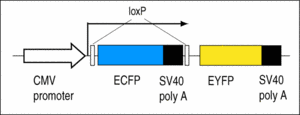
Y2: Contains the Cre reporter CMV:ECFP(loxP)EYFP.
The larvae show blue fluorescence, which changes to yellow fluorescence when exposed to Cre recombinase (Ryffel et al., 2003).
GpL8:
Contains the Cre reporter CMV:ECFP(loxP)LacZ.Larvae exhibit green fluorescence that changes to ß-galactosidase activity upon exposure to Cre recombinase, which can be visualized by LacZ staining (Waldner et al., 2006).
Tom3:
This strain contains a CMV-driven red fluorescent protein (tdTomato, kindly provided by Shaner et al., 2004). Adults of this strain show ubiquitous and homogeneous expression of the red fluorescent protein (Fig. 1), indicating that the CMV promoter can also be used to express transgenes at later developmental stages (Waldner et al., 2006).
The figure shows an adult frog of the CMV:tdTomato strain tom3.
A: Head illuminated for red fluorescence.
B and C: Prepared mesonephros of a tom3 animal (right) or a control animal (left) in normal light (B) or red fluorescence (C).
(b) EFFECTOR STRAINS
328del-4:
Contains the Cre/FLP effector CMV:ECFP(loxP)(FRT)HNF1BP328L329del.The larvae show blue fluorescence and express the human HNF1B mutation P328L329 after Cre recombinase action (Roose et al., 2009).
328del-3:
Contains the Cre/FLP effector CMV:ECFP(loxP)(FRT)HNF1BP328L329del and is a line independent of 328del-4 with very similar properties. The larvae show blue fluorescence and express the human HNF1B mutant P328L329 after Cre recombinase action (Roose et al., 2009).
A263ins-6:
Contains the Cre/FLP effector CMV:ECFP(loxP)(FRT)HNF1BA263insGG. The larvae show blue fluorescence and express the human HNF1B mutation A263insGG after Cre recombinase action (Sauert et al., 2011).
(c) RECOMBINASE STRAINS
A7:
Contains the CAR-Cre construct encoding the coding sequence of Cre recombinase under the control of the cardiac actin promoter. Cre activity was demonstrated by crossing with the reporter strains C5, Y2 and GpL8 (Waldner et al., 2006).
HSPCre1:
This strain contains a Cre recombinase under the heat shock promoter HSP70. F1 animals of this strain crossed with the C5 reporter strain show ubiquitous induction of the EYFP reporter upon heat shock. DNA recombination was observed as early as one hour after the start of heat shock treatment (Roose et al., 2009).
HSPCre1;C5:
These frogs originate from a cross between HSPCre1 and C5. They were heat shocked during embryogenesis and the yellow larvae (activated EYFP) were reared.
HSPCre13:
This strain contains a Cre recombinase under the heat shock promoter HSP70. Since this strain contains a linked CMV-driven gene encoding the red fluorescent protein tdTomato, progeny with Cre recombinase can be easily identified. F1 animals of this strain crossed with the C5 reporter strain show ubiquitous induction of the EYFP reporter upon heat shock (Roose et al., 2009; Sauert et al., 2011).
Reference list
Roose,M., Sauert,K., Turan,G., Solomentsew,N., Werdien,D., Pramanik,K., Senkel,S., Ryffel,G.U., and Waldner,C. (2009). Heat-shock inducible Cre strains to study organogenesis in transgenic Xenopus laevis. Transgenic Res. 18, 595-605.
Ryffel,G.U., Werdien,D., Turan,G., Gerhards,A., Goosses,S., and Senkel,S. (2003). Tagging muscle cell lineages in development and tail regeneration using Cre recombinase in transgenic Xenopus. Nucleic Acids Res. 31, e44.
Sauert,K., Kahnert,S., Roose,M., Gull,M., Brändli,A.W., Ryffel,G.U., and Waldner,C. (2011). Heat-shock mediated overexpression of HNF1b mutations has differential effects on gene expression in the Xenopus pronephric kidney. PLoS. ONE. submitted.
Waldner,C., Roose,M., and Ryffel,G.U. (2009). Red fluorescent Xenopus laevis: a new tool for grafting analysis. BMC. Dev. Biol. 9, 37.
Waldner,C., Sakamaki,K., Ueno,N., Turan,G., and Ryffel,G.U. (2006). Transgenic Xenopus laevis strain expressing cre recombinase in muscle cells. Dev. Dyn. 235, 2220-2228.

Univ.-Prof. Dr.
Gerhart U. Ryffel
Im Ruhestand
(Retired)